
初期エンドソームの膜骨格
中川 裕之
九州工業大学
前シナプス膜に融合したシナプス小胞膜は、エンドサイトーシスによって再び小胞として速やかに細胞内に取り込まれる。 この取り込み機構は、ほとんど全ての真核細胞で観察されるクラスリン依存的エンドサイトーシス機構と同一である。 細胞膜からちぎれた小胞は初期エンドソームへと輸送され融合する。 この過程にアクチンが関係していることが報告されているが、その具体的な役割についてはまったく分かっていない。 そこでアクチン分子と初期エンドソームの細胞内局在を詳細に観察したところ、 アクチン分子が初期エンドソームの中でもソーティングエンドソームに繊維として存在することが分かった。 この結果は、アクチン繊維がソーティングエンドソームの膜骨格としてエンドソームの形や小胞との融合の制御に関わっていることを示唆する。
[HOME]1998年4月23日
海馬CA3領域のLTPと自発活動
中島 孔志
九州工業大学
海馬CA3錐体細胞には大きく2種類の興奮性シナプスがある。一つは歯状回の顆粒細胞からCA3錐体細胞へのびる苔状線維を介した苔状線維シナプス、他方はCA3 領域内の錐体細胞同士が相互に結合する相互シナプスである。これら2種のシナプスはそれぞれ高頻度刺激によりシナプス伝達効率の長期増強を引き起こすことが知られている。しかし、それらのシナプス伝達効率の変化とCA3領域の自発活動との関係はわかっていない。海馬での情報処理機構解明の上でこれらの関係は大変興味深い。
本研究ではCA3領域が10-20mmとかなり長く得られ、複数のラメラ構造を含むラット海馬縦断面スライスと16chマルチ電極を用い、苔状線維の高頻度刺激によって生じたLTPの空間分布とCA3錐体細胞層の自発活動との関係について調べた。その結果、苔状線維を高頻度刺激した結果、苔状線維シナプスには局所的にLTPが生じたがCA3錐体細胞層には顕著な自発活動は生じなかった。苔状線維にLTPがおきている状態で、さらに苔状線維を高頻度刺激するとCA3錐体細胞層で自発活動としててんかん様波が観測された。苔状線維シナプスのLTPによってCA3錐体細胞への入力が強まり、相互シナプスのLTPがかなりの広範囲で起き、その結果てんかん様波が生じたこと示している。
1998年4月30日
神経回路網の確率共鳴とその記憶想起モデルへの応用の可能性
林 初男
九州工業大学
双安定または単安定状態が存在する非線形系では,雑音が存在することにより信号対雑音比が向上するという現象が見られ,確率共鳴(stochastic resonance)と呼ばれている.ニューロンも確率共鳴を起こすことが示されており,末梢の感覚受容器が雑音に埋もれた信号を検出する機構の一つであると考えられている.前前回の担当では確率共鳴のレビューをし,前回の担当では一個のニューロンの確率共鳴について報告した.今回は,神経回路網の確率共鳴を海馬CA1神経回路網モデルを用いて示し,さらに,確率共鳴現象の記憶想起モデルへの応用の可能性を示す.
[HOME]1998年5月21日
Modeling and analysis of spatio-temporal change in [Ca2+]i in a retinal rod outer segment.
K. Ichikawa. Neuroscience Research 25, 137-144, 1996.
八木 哲也
九州工業大学
今週は、細胞内カルシウムの動態に関する計算機シミレーションの論文を紹介します。
[HOME]1998年5月28日
明順応信号としての一酸化窒素(NO)とH1水平細胞の短波長性入力
古川 徹生
九州工業大学
気体性の伝達物質であるNOが網膜神経系に対して明順応と同じ効果を示すことがわかってきた.コイ網膜のH1水平細胞は赤錐体から主入力を受けるが,同時に短波長性入力によって光応答特性が修飾される.この短波長性入力は明順応時にのみ存在し,暗順応時には不活性化する.NOは明順応と同様にこの短波長性入力を活性化する働きがある.われわれは以上のことを (1) 波長感度特性 (2) 光誘起性入力抵抗変化 (3) 受容野 の3点から確認してきた.今回の発表では今までの結果を総括し,またH1水平細胞におけるNOおよび短波長性の視覚情報処理における機能について検討したい.
[HOME]1998年6月11日
カエルのイスミ核と視蓋
星野 徳明
九州工業大学
カエルの視覚系は、一方の網膜からの入力が対側視蓋に投射する完全交叉となっ ていますが、視蓋の前方から3分の1 程度の部分に両眼視の機能があることが 知られています。ただし、同側の網膜から直接の入力はなく、いったん対側の イスミ核を経由し、イスミ-視蓋経路から同側の入力を得ています。今回はそ の両眼視に深くかかわるイスミ核の機能などについて発表します。
[HOME]1998年6月18日
Binding問題について
安井 湘三
九州工業大学
今週は私が担当ということで、何をネタにしようかと迷いましたが、binding
問題を自分の番がまわってきたときに続きもので紹介していこうと決めました。
最近急速に注目されてきた分野で、これが解明されなければ脳が理解されるこ
とはないと考えます。今回は、まずBinding 問題とはそもそも何かということ、
つまり「問題の問題」を(2)のサーベイ論文などを参考に皆さんと考えて行き
たいと思います。(1)は今から読んで勉強するところですが、これを選んだ理
由は、これが比較的最近のNatureに載っていたからというだけです。良い論文
であることを期待しております。もしそうでなかったら今回はご容赦ください。
次回の担当のときからは、もっと私なりの計画性をもってBinding問題に関し
た具体的な論文を紹介していくつもりでいます。
(1) Luck,S.J. & Edward K.V.
The capacity of visual working memory for features and conjunctiuons. Nature 390:279-281 (1997)
(2)Treisman,A., The binding problem. Curr. Opinion Neurobiol.,6:171-178 (1996).
1998年6月25日
八木 哲也
九州工業大学
最近、生体神経回路の並列情報処理機構を直接集積回路化する動きがあります。 特に、現在のノイマン型直列コンピュータがもっとも不得手とする感覚情報処 理を実行するアナログチップ(例えばビジョンチップ)は、生体神経回路の計算 原理を構成的に研究するためのパラダイムとしても、ロボティクス等の工学な 応用面からも将来の可能性が注目されています。しかしながらこのシステムの 究極的な意義は、脳神経システムの一部を代行させることにあるのではないか と考えています。この気の遠くなるような目的の第一歩として、いかに実際の 神経と集積回路の間で情報の伝達をおこなうかという問題を解決しなければな りません。今回のセミナでは、神経からの電気信号を基盤電極などで受ける手 法について以下の論文を中心に紹介します。
A Neuron-Silicon Junction: A Retzius Cell of the Leech on an Insulated-Gate Field-Effect Transistor.[HOME]
Science, vol.252, 1290-1293, 1991.
by P. Fromherz, et. al.
その他の関係論文
1998年7月9日
電位依存性Caチャネルの薬理学
吉井 清哲
九州工業大学
細胞内のCaの濃度を細胞外の1/10000程度に減少させることで、動物はCaを細 胞内メッセンジャーとして利用している。細胞外からの主要なCa流入経路を形 成する電位依存性Caチャネルの薬理学的性質について、Campbellらの総説を紹 介する。
[HOME]1998年9月10日
ラット上丘におけるθ波関連細胞
夏目季代久
九州工業大学
ウレタン麻酔下ラットにおけるθ波及びθ波関連細胞を感覚運動統合過程に関 係している上丘領域より調べた。結果、1)上丘においてθ波が観察された。 2)Tonicなθ-on細胞、θ-off細胞が見つかった。3)θ-on 細胞の平均周波 数は、θ波誘導と共に増加した。4)θ-off細胞の周波数は、θ波誘導と共に 減少した。5)上丘は、表層から、浅層、中間層、深層に分類され、θ-on細 胞は各層均一に分布していたが、θ-off細胞は主に上丘の中間層、深層に分布 していた。海馬におけるθ-off細胞は、海馬θ波誘導に関係しているので、上 丘の中間層、深層が上丘におけるθ波発生に関与していると考えられる。また 感覚運動統合過程は、上丘深層で行なわれていると考えられているので、θ波 が、その情報処理過程に何らかの関与をしているかも知れない。
[HOME]1998年9月17日
遺伝的プログラミングとその適用例
岡本正宏
九州工業大学
進化論的な考えに基づいてデータを操作して最適化問題や学習
推論を扱う方法の1つに、”遺伝的アルゴリズム”がある。
この方法の手順は次のようにまとめられる。
(1)対象とする問題の解を染色体上にコード化する。一方、この遺伝的アルゴリズムと同じく自然淘汰と遺伝子変異の仕組みを応用 したものに、遺伝的プログラミングがある。遺伝的アルゴリズムとの違いは、 遺伝的プログラミングは、進化の機構を用いてプログラムを自動生成すること を目的としている。つまり、この方法を用いると、目的のプログラムの内部構 造や処理手順について明示的に設計することなく、プログラムを生成すること が可能になる。セミナーでは、遺伝的アルゴリズムと対比させながら、遺伝的 プログラミングを概説し、その適用例を紹介する。 [HOME]
(2)染色体を複数個用意して集団を形成する。(初期集団ははランダムに生成される。)
(3)集団に対して淘汰をかけ、優れた個体を選択する。
(4)遺伝子操作(交差、突然変異)により、新しい解を探索する。
(5)選択、遺伝子操作を世代というサイクルでくりかえし、より優れた解を探索する。
1998年10月8日
CA3神経回路網モデルの時空間放電パターンとその電場リズム
立野 勝巳
九州工業大学
海馬で観測されるリズムは機能的な動作との関わりが深く、興味深い研究対象
です。しかしながら、それらのリズムに対応する神経回路網の時空間的な性質
は十分に明らかにされておりません。本研究では4種類の結合パターンでCA3
神経回路網を構成し、その時空間活動とそれに対応するリズムを調べました。
その結果、興奮性結合と抑制性結合のバランスに依存して、神経回路網は周期
的なリズムと不規則なリズムを示しました。本研究では、それらのリズムの空
間的な相互相関関数を求め、空間的な活動を評価しました。リズム生成の違い
は、同期して活動するニューロン集団のサイズの変化、および伝播により生じ
ることが分かりました。
1998年10月15日
李 麗明
This is Liming Li in Yagi's Lab.. This time I would like to tell you the abstract of my presentation on this Thursday seminar.It is mainly about the visual prosthesis and our preliminary attempts to establish the multineuronal stimulating-recording system.
[HOME]1998年10月29日
中川 秀樹
九州工業大学
Elementary computation of object approach by a wide-field visual neuron
N. Hatsopoulos, F. Gabbiani and G. Laurent Science 270 (1995)
Local circuit for the computation of object approach by an identified visual neurons in the locust
F. C. Rind and P. J. Simmons J. Comp. Neurol. 395 (1998)
Collision-avoidance: nature's many solutions
G. Laurent and F. Gabbiani Nature Neurosci. 1(4) (1998)
Computation of different optical variables of looming objects in pegion nucleus rotundus neurons
H. Sun and B. J. Frost Nature Neurosci. 1(4) (1998)
ハトは接近してくる物体の持つ異なる3つのパラメータを符号化する神経を持っ
ています。またそのうちの1つは、何と、系統的にかけ離れた動物であるバッ
タにも見られます。以上の論文を題材に、動物が、近づいてきてついにはぶつ
かってしまう物体をいかにして検出することができるのか。自然の用意したそ
の巧妙な手口(その神経機構)についての興味深い報告をご紹介したいと思い
ます。
1998年11月12日
細胞膜骨格の電子顕微鏡による観察
中川 裕之
九州工業大学
真核細胞の細胞膜には膜骨格と呼ばれる構造が存在し、細胞質側から膜を保持 していると考えられている。この膜骨格の研究は、簡単に細胞膜を単離できる 哺乳類の赤血球膜をモデルとして行われてきた。赤血球はそれ自身で運動する ことのない細胞だが、赤血球以外のほとんどの真核細胞はエクソ/エンドサイ トーシスや細胞運動によってつねに細胞の形態を変化させている。このような 形態の変化と細胞膜骨格との関わりは現在まで明らかではない。そこで細胞膜 骨格を細胞質側から電子顕微鏡で観察する方法を開発し、培養繊維芽細胞の膜 骨格を観察した。
[HOME]1998年11月19日
海馬CA3縦断面スライスにおける錐体細胞相互結合シナプスのLTPと自発性リズム
中島 孔志
九州工業大学
ラット海馬CA3領域は縦断面方向に1cm以上あり,いくつかのラメラ構造が機能
分担していると考えられている.海馬内のシナプスは大きなシナプス可塑性を
有し,シナプスの伝達効率が海馬の状態によってさまざまな変化を起こす.そ
のためラメラ構造は海馬の状態によってさまざまに変化するのではないかと考
えられる.また,このような海馬縦断面方向の空間的な特徴が海馬の情報処理
過程を左右しているのではないかと考えられる.海馬における情報処理過程の
理解するためには海馬縦断面方向の時空間的な電気生理学的性質を理解するこ
とが必要である.
我々はスライス試料で空間的に同時観測可能なマルチ電極を開発した.その
マルチ電極とラット海馬縦断面スライスを用いて,苔状線維高頻度刺激により
生じた単シナプスLTPが局所的に生成されることを報告した.しかし,単シナ
プスLTPが生成されても自発性のリズムは観測されず,単シナプスLTP生成後再
度の高頻度刺激により初めて自発性てんかん様バーストが全域で生じた.これ
らの結果をふまえ,高頻度刺激後に1分間間隔でテスト刺激を加えそれによっ
て生じる単および多シナプスLTPと自発性リズムの関係について報告する.ま
た,これまでのスライスを用いた結果より示唆される海馬CA3領域の全体像に
ついてのまとめを報告する.
1998年11月26日
中隔と後部視床下部同時刺激によるラット海馬θリズム及び輪廻し行動の変化
夏目季代久
九州工業大学
海馬で観測される脳波θリズム(HPθ)は、上行性脳幹神経系の活動によって 生じる事が分かっている。上行性脳幹神経系は、脚橋被蓋核から後部視床下部 (PH)、中隔(MS)を通って海馬、内嗅領に至る経路である。Blandらによれば、 PH刺激を行なうと、刺激強度に応じて、1)HPθの周波数が増加する事、2) ラットの輪廻し速度が増加する事が観測された。またそれらの結果がMSの局所 麻酔により阻害される事が明らかになった。従って、PHは、MSを経由した経路 でHPθの周波数及び輪廻し速度を制御しうる可能性が示唆された。また以前の 結果では、HPθの周波数はMS刺激によっても影響される。MSの5Hz刺激により 5HzのHPθ、10Hz刺激により10HzのHPθが出現する。Darrenらによる昨年の Neurosci. Meetingでの発表によれば、急性ウレタン麻酔下ラットにおいて、 MSとPHの同時刺激を行なうとHPθの周波数はMS刺激のみに従う。つまり、PH刺 激によって、いかなる周波数が出ていても、MSを5Hzで刺激すれば5HzのHPθが、 10Hzで刺激すれば10HzのHPθの出現が観察される。私たちは、この刺激実験を 慢性電極を埋め込んだラットで行なった。また同時に刺激によるラット輪廻し 行動への影響について観察した。その結果について、セミナー当日は発表しま す。まだまだデータは少ないですが、面白い結果が出ましたので、皆さんの御 意見をお聞かせ下さい。
[HOME]1998年12月17日
ニューロン活動の同期化と低次元カオス -てんかん脳波はカオスか-
林 初男
九州工業大学
1985年頃から始まった脳のカオス活動の探究の結果,非常にたくさんの論文が
提出されたにも関わらず,10年近くもその証拠は示されなかった.2,3年前,
我々は海馬CA3と新皮質体性感覚野の周期刺激に対するカオス応答の証拠を示
すことができたが,なぜ首尾よくカオス応答を観測できたのか,その理由は当
時よく分からなかった.結論的に言えば,周期刺激によって空間的自由度が減
少したにも関わらず時間的複雑さが残ったからである.このようにニューロン
活動の同期化に伴って脳のカオス的活動が表面化するのであれば,脳のカオス
的活動を観測できるいくつかの場合を考えることができる: (1)入力信号によ
るニューロン活動の短期的同期化の促進,(2)LTPのようなシナプスの可塑的変
化に伴うニューロン活動の長期的同期化の促進,(3)てんかんのような病的な
脳の活動などである.
今回は,まず,ニューロン活動の同期化と脳のカオス的活動という観点から,
今までの海馬神経回路網モデルの結果やラットの海馬および体性感覚野の実験
結果をざっと見直す.そのあと,今年の秋オランダのてんかん研究所で明らか
にできた,ヒトのてんかん脳波のカオス的活動について述べる.
1999年1月14日
神経細胞における細胞内Ca2+ストアの機能について
林田 祐樹
九州工業大学
近年、神経細胞内に存在するCa2+ストアが、細胞膜上に存在する伝達物質駆動 型チャネルや電位依存性チャネルのコンダクタンスを調節するうえで重要な役 割を持っていることが示されています。例えば網膜の神経節細胞では、(1)リ アノジン感受性Ca2+ストアからのCa2+放出により、GABA投与により誘発される Cl電流が抑制されることや(Akopian et al., 1998)、(2)グルタミン酸による 電位依存性Ca2+電流の抑制にリアノジン及びIP3感受性Ca2+ストアが関与する こと(Shen & Slaughter, 1998)が報告されています。また小脳では、プルキニェ 細胞のグルタミン酸応答のLTDにCa2+ストアからのCa2+放出が必要であること (Kohda et al., 1995)、顆粒細胞の電位依存性Ca2+チャネルの活性をリアノジ ン感受性Ca2+ストアが調節すること(Chavis et al., 1996)などが報告されて います。今回の発表では、以上の論文を簡単に紹介するとともに、結論はほと んど未だ出ていませんが、網膜水平細胞で行なったCa2+ストアに関する私の実 験結果について紹介させて頂きます。
[HOME]1999年1月28日
違いのわかる抗てんかん薬チアガビン
笛田 由紀子
産業医科大学
後シナプス膜電位を過分極するという抑制性シナプス伝達機構の減弱は、神経
細胞群の過剰興奮の原因と考えられている(てんかんのGABA仮説)。我々は、
てんかんの過剰興奮の原因機構を調べるために、自然発症のてんかんモデルEl
マウスの海馬におけるGABA性抑制を電気生理学的に調べてきた。そして、GABA
性抑制の変化は、1)発作感受性の増加に正相関する変化(growth-dependent)
と、2)発作感受性のない、若いElマウスにすでに見られる変化
(growth-independent) に分類された。Elマウス海馬CA1のGABAシナプスの伝
達効率の減少はGABA放出量の減少によるという報告に引き続き、再取り込みを
阻害してシナプス間隙の有効GABA量を増やして発作を抑制できるかを実験した。
チアガビンは、血液-脳関門を通過するGABAトランスポーター阻害剤として
Novo Nordisk (Denmark)で開発され、現在日本でも臨床試験中である。今回の
セミナーでは、Elマウスの海馬スライスの実験で報告してきた興奮性の変化
に対する、チアガビンの作用を報告する。この発表の主眼は、単に抗てんかん
薬の効果を報告するのではなく、チアガビンがシナプス環境選択的に作用する
という点、すなわち、「違いのわかる抗てんかん薬」であるという点にある。
「シナプス環境選択的」という、あえて、曖昧な表現をしましたが、ここのと
ころについて議論をしていただければと思います。
1999年3月11日
網膜神経系における順応信号としての一酸化窒素
古川 徹生
九州工業大学
前回のセミナーで,コイ網膜のH1型水平細胞は赤錐体から主入力を受けつつも,短波長感受性錐体(青 and/or 緑錐体)から符号反転型の入力を受けていること,そしてこの入力は明順応時にのみ活性化し,一酸化窒素(NO)がその活性化に関与することをお話した.すなわち以下の構図が成り立つ.
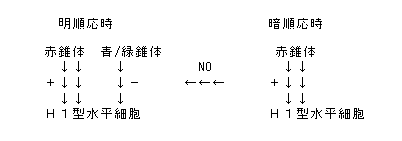
前回のセミナーでは,波長感度特性の順応に伴う変化から上記の構図が成り立つことを示した.ただしこれだけでは短波長感受性錐体からH1水平細胞への入力経路が直接のシナプス結合によるものか,第三の細胞を介した間接的なものかはわからない.
今回は,H1の受容野の大きさが光刺激の波長によって変化すること,そして順応及びNOによって受容野の波長依存性が変化することから,上図の構図が成り立つことを示す.
さらに短波長感受性錐体からH1水平細胞への入力経路を調べるため,光刺激に伴うH1水平細胞の入力抵抗変化を測定した.この「光誘起性入力抵抗変化」も受容野同様に刺激光に依存した波長特性を持ち,順応とNOによって変化した.この結果は,短波長感受性錐体からH1への入力が直接的なものであることを示唆する.